M. Barry Sterman, PhD Professor Emeritus, Departments of Neurobiology and Biobehavioral Psychiatry, Geffen School of Medicine, University of California, Los Angeles, CA
Correspondence: M.B. Sterman, PhD, 9773 Blantyre Drive, Beverly Hills, CA 90210; msterman@ucla.edu
Dr. Sterman reported that he has intellectual property rights and ownership in Sterman-Kaiser Imaging Laboratory, for which he also is a member of the board of directors.
ABSTRACT
This review traces the application of electroencephalographic (EEG) operant conditioning, or biofeedback, from animal research to its emergence as an alternative treatment for the major types of seizure disorder. Initial animal studies focusing on brain mechanisms that mediate learned behavioral inhibition revealed a uniquely correlated 12- to 15-Hz EEG rhythm localized to sensorimotor cortex. We labeled this the sensorimotor rhythm, or SMR. The similarity of the SMR to the known EEG spindle pattern during quiet sleep led to the novel idea of attempting to increase the SMR using EEG operant conditioning. The hypothesis was that this might produce a corresponding increase in sleep spindle activity, thus establishing a common EEG marker for the state of motor inhibition. Results supported this hypothesis but led also to the accidental discovery of an anticonvulsant effect on drug-induced seizures in cats and monkeys. Continuing animal studies identified a pattern of neurophysiologic responses correlated with the SMR in primary motor pathways. These and other findings were indicative of reduced motor excitability. Simultaneously, we undertook studies in human epileptic subjects that documented a significant reduction in seizure incidence and severity, together with EEG pattern normalization. This work expanded internationally, resulting in numerous well-controlled group and single-case studies summarized in recent meta-analyses. Exciting new findings in functional neuroimaging/EEG correlation studies provide a rational model for the basis of these clinical effects. In recognition of the diversity of clinical applications of EEG biofeedback and the complexity of seizure disorders, this review also details specific methods used in our EEG biofeedback program.
The attempt to alter electroencephalographic (EEG) frequency/amplitude patterns and their underlying brain mechanisms using contingent operant conditioning methods is today referred to variously as EEG biofeedback, neurofeedback, or neurotherapy. This article traces the history of the clinical application of EEG operant conditioning from empirical animal investigations to its emergence as a treatment option for major seizure types. In light of the diversity of the clinical applications of this method in general, and the complexity of seizure disorders in particular, I also present an overview of specific methods used in our EEG biofeedback program.
Figure 1. A carefully documented 6-year seizure data log from an adult female subject (aged 23 years at the start of the log) with nocturnal tonic-clonic seizures, often with incontinence.2 The log starts 1 year before initiation of electroencephalographic (EEG) feedback training (“Pre-SMR”), continues through 2.5 years of twice-weekly EEG training sessions (“Post-SMR”), and continues through 2.5 years after wi thdrawal from this training (“Withdrawal”). Training consisted of auditory and visual reward for increased 12- to 15-Hz EEG activity over the left sensorimotor cortex, which has been labeled the sensorimotor rhythm (SMR). Medications were held constant during training and adjusted downward after withdrawal from training. In 1977 this patient was issued a California driver’s license.
This application was officially added to the broader field of biofeedback with the publication of a 1972 paper by Sterman and Friar titled, “Suppression of seizures in an epileptic following sensorimotor EEG feedback training.” 1 In this paper we documented a sustained and progressive reduction of generalized nocturnal tonic-clonic seizures in a 23-year-old female epileptic with a 7-year history of frequent and medically refractory seizures of unknown origin. The patient’s clinical EEG showed left sensorimotor cortex spikes and slow 5- to 7-Hz activity. Seizure reduction occurred in response to an experimental course of EEG operant conditioning aimed at increasing 12- to 15-Hz EEG activity in the left sensorimotor cortex while suppressing slower activity at this same site. The 12- to 15-Hz EEG rhythm was discovered in animal research and labeled as the sensorimotor rhythm (SMR). Although the patient had previously been worked up and treated unsuccessfully with anticonvulsant medications at several prestigious medical institutions, over the course of 2.5 years of twice-weekly EEG feedback training sessions she became essentially seizure free (Figure 1)2 and was ultimately issued a California driver’s license.
BACKDROP TO THE CLINICAL APPLICATION: KEY ANIMAL STUDIES
Figure reprinted from Sterman et al (Science 1970; 167:1146–1148).3
Figure 2. Bipolar electroencephalographic (EEG) samples from sensorimotor and parietal cortex in the cat during quiet (motionless) wakefulness (left) and quiet (non-REM) sleep (right). Both states are associated with bursts of 12- to 15-Hz EEG rhythmic activity in sensorimotor cortex. During sleep these bursts are higher in amplitude and associated with slower rhythmic patterns in parietal cortex.
The above landmark study was predicated on the observation of a discrete 11- to 19-Hz EEG rhythmic pattern in cats, which occurred intermittently over the sensorimotor cortex during behavioral quiescence. When animals were trained to suppress a learned bar-press response for food if a tone was sounded in the chamber, a 12- to 15-Hz version of this EEG pattern always accompanied inhibition of the bar-press response. If animals later fell asleep, a similar rhythmic EEG pattern, known as the sleep spindle, was localized to the same cortical area at the same frequency (Figure 2).3 Our interest at the time was in the neurophysiological control of sleep. Because both of these patterns occurred uniquely in the absence of movement, we sought to determine if the underlying neural mechanisms were related.
To accomplish this, we attempted to facilitate the SMR during wakefulness using an operant conditioning paradigm with a liquid food reward, and then study any resulting changes in sleep spindle activity and sleep structure. Necessary quality controls included alternate training to suppress this rhythm and a counterbalanced design employing two separate groups of cats. Six weeks of three training sessions per week to satiation led to profound and differential changes in sleep EEG and sleep architecture. SMR training, whether it preceded or followed suppression training, led to a significant increase in EEG sleep spindle density, as well as a significant reduction in sleep period fragmentation due to arousals. No changes occurred in the control condition.3
A more profound finding in the cat
Figure modified from Sterman.5
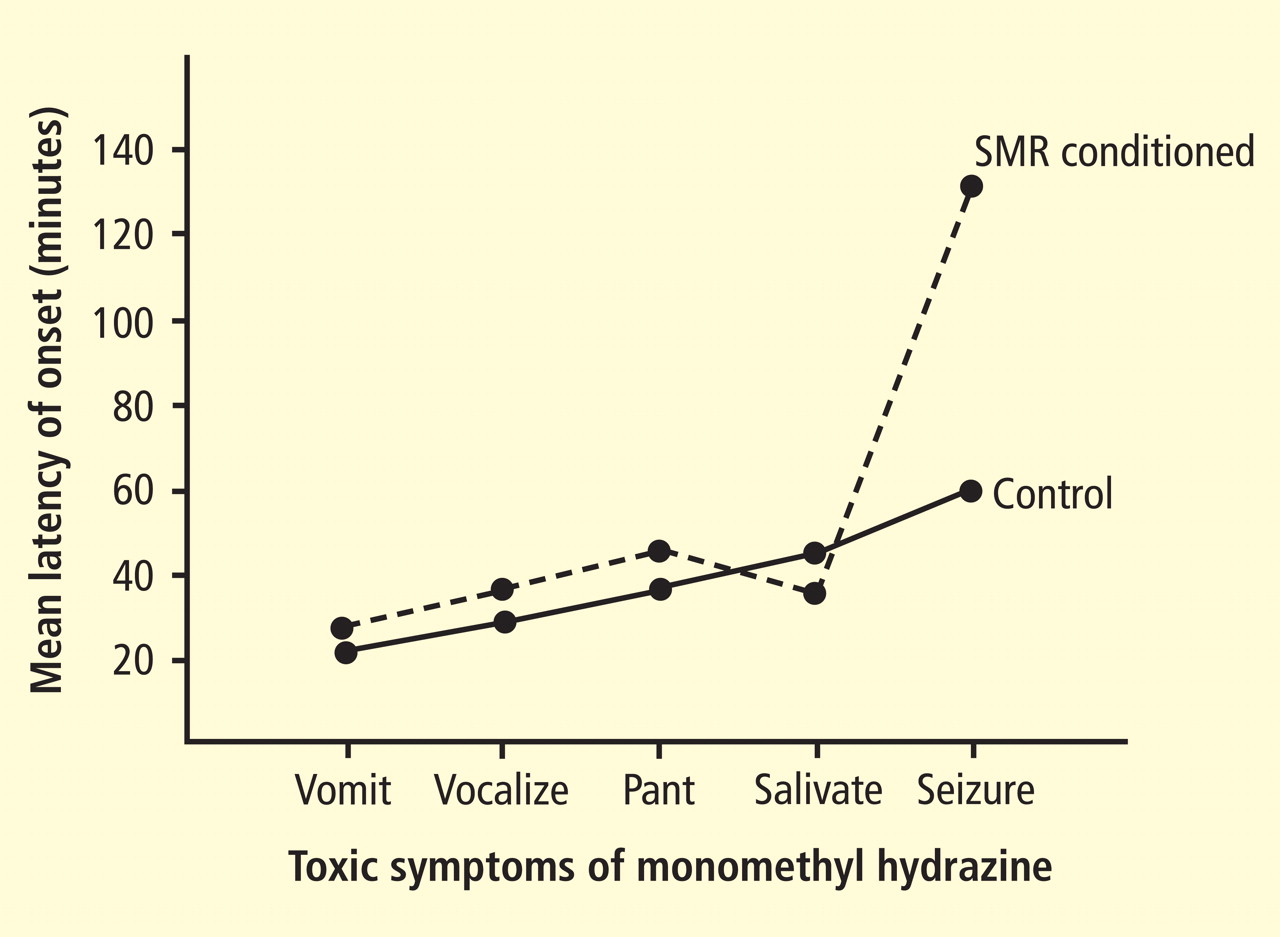
Figure 3. The sequence of prodromal events preceding generalized convulsions in two groups of 10 cats, all of which were injected intra-abdominally with 9 mg/kg of GABA-depleting monomethyl hydrazine. One group (dashed tracing) had received 6 weeks of electroencephalographic feedback training for sensorimotor rhythm (SMR) enhancement with food reward (see text). The two groups did not differ statistically in the latency to prodromal symptoms. All control animals seized reliably at approximately 60 minutes, as had been previously documented. In contrast, the SMR-trained group had a significantly prolonged mean latency to seizures (130 minutes), and several did not seize within the 4-hour test period.
As interesting as this finding was, the most profound outcome of the study emerged later. A different cat study under way in our laboratory, funded by the US Air Force, was seeking to determine the effects on behavior of low-dose exposure to monomethyl hydrazine (MMH).4 This compound is a highly toxic component of the liquid rocket fuel used for launching virtually all space vehicles. Significant MMH exposure via any route causes pro-found nausea and gradual onset of convulsions, which are lethal at adequate doses. The mechanism for this effect was ultimately determined to be a disruption of the synthesis of gamma-aminobutyric acid, the primary inhibitory neurotransmitter in the central nervous system. We were investigating the effects of low-dose exposure to determine the possible disruption of cognitive functions such exposure might cause in flight crews. Our first objective for studies in cats was to establish the doseresponse curve for convulsive effects in that species. We had succeeded in determining a curve showing that 9 mg/kg of MMH was the threshold dose for reliably producing nonlethal convulsions after a prodrome of approximately 40 to 67 minutes. This prodrome consisted of a sequence of reliable autonomic and behavioral events. When data from animals provided with SMR operant conditioning as the final training procedure were added to this curve, the same prodrome was observed but there were no seizures at 60 minutes. Instead, the latency to seizures was delayed to a range of 80 to 220 minutes, and several animals failed to seize at all.4 A subsequent systematic study of this effect with animals as their own controls in a counterbalanced design confirmed this effect (Figure 3).5 This finding then led to the test in the human epileptic subject described above.1