Pinacidil induces vascular dilation and hyperemia in vivo and does not impact biophysical properties of neurons and astrocytes in vitro





ABSTRACT
Vascular and neural systems are highly interdependent, as evidenced by the wealth of intrinsic modulators shared by the two systems. We tested the hypothesis that pinacidil, a selective agonist for the SUR2B receptor found on smooth muscles, could serve as an independent means of inducing vasodilation and increased local blood volume to emulate functional hyperemia. Application of pinacidil induced vasodilation and increased blood volume in the in vivo neocortex in anesthetized rats and awake mice. Direct application of this agent to the in vitro neocortical slice had no direct impact on biophysical properties of neurons or astrocytes assessed with whole-cell recording. These findings suggest that pinacidil provides an effective and selective means for inducing hyperemia in vivo, and may provide a useful tool in directly testing the impact of hemodynamics on neural activity, as recently predicted by the hemo-neural hypothesis.
Pinacidil administration in vivo
Pinacidil is hydrophobic and was therefore dissolved in ethanol at approximately 12 mg/kg and then diluted 1:100 in ACSF to achieve a 400-μM solution in 1% ethanol. Stock solutions were stored at −20°C and diluted in fresh ACSF for each experiment. For each run, the cortex was imaged for 3 minutes to establish baseline. For the pressurized rat chamber, at the end of the baseline period, 0.1 to 0.3 mL of 400 μM pinacidil in 1% ethanol in ACSF or saline would be pumped into the 0.3-mL chamber (taking about 1 second). Simultaneously, an equivalent volume was drawn out to balance pressure by a push-pull pump with access through two of the resealable rubber ports.
Pinacidil administration in vitro
Coronal slices were prepared from Sprague-Dawley rats at postnatal day 14 to 40 and maintained in a submersion chamber at 27°C for recording. Solutions were prepared in ACSF: 125 mM NaCl, 2.5 mM KCl, 1 mM MgCl2, 1.25 mM NaH2PO4, 2 mM CaCl2, 25 mM NaHCO3, and 25 mM d-glucose). The applied solutions were 1% ethanol, ~400 μM pinacidil in 1% ethanol, and ACSF, all perfused with 5% CO2 in 95% O2 (carbogen).
For astrocyte recordings, slices were incubated immediately after cutting for 20 minutes in ACSF containing 50 μM sulforhodamine 101 (SR101), a water-soluble fluorescent dye specifically taken up by astrocytes.19 The slices were then allowed to rest for an hour before recording as usual. For fluorescence, the light source was a 100-W mercury arc lamp, with excitation and barrier filters and dichroic mirrors tailored to the spectral characteristics of SR101 (excitation ~586 nm, emission ~605 nm)
Slices were imaged under differential interference contrast optics with infrared illumination. Cells in layers 2/3 and in the same field and plane of view as a blood vessel greater than 20 μm in diameter were targeted, and vessel expansion was monitored during intracellular recording at approximately 4 Hz with a cooled charge-coupled device camera (Retiga EX, QImaging, Surrey, BC, Canada) connected to the microscope via a parfocal C-mount. This configuration also enabled imaging of neurons and astrocytes in the slice.
Drug/control solutions were switched every 90 to 180 seconds. Recording pipettes were filled with 120 mM KGlu, 10 mM NaCl, 20 mM KCl, 10 mM HEPES, 2mM Mg-ATP, 0.3 mM Na-GTP, 0.5 mM EGTA, and 0.3% to 1% biocytin (wt/vol) for subsequent visualization of the neurons.
RESULTS
Pinacidil induces vasodilation in anesthetized rats and awake mice
Figure 1A shows the cortical surface over SI. The green bisection line over the MCA shows the point of sampling for the darkness plot in the rectangular box in Figure 1B. The width of this dark band is the width of the MCA over time, showing expansion after pinacidil addition (gray bar). The point at which dilation is observed corresponds to a darkening in a parenchymal region (red triangle in Figure 1A) and over the MCA and surrounding cortex (purple rectangle in Figure 1A, reflecting expansion). Vasodilation and parenchymal signal increases were consistently observed on the first trial in all experiments (4 first runs from 4 anesthetized rats, Figure 1C and 1D). Dilation began less than 10 seconds after drug arrival, with maximal dilation and parenchymal darkening at an approximately 50- to 60-second latency. Ethanol in a 1:100 solution with ACSF under the same conditions evoked a nonsignificant reduction in vessel diameter and no change in parenchymal darkening. Following the first presentation, subsequent pinacidil effects were less reliable.
As shown in Figure 1E and 1F, the hemodynamic impact of pinacidil in anesthetized rats was replicated in awake, head-posted mice (2 mice, 2 runs each). Presentation of 220 or 440 μM pinacidil evoked comparable mean increases in arterial diameter (peak diameter increase of ~20%) and parenchymal darkening (peak increase of ~2%), effects that were repeatable within subjects in a single session (N = 2).
Pinacidil does not have direct effect on nonvascular tissue
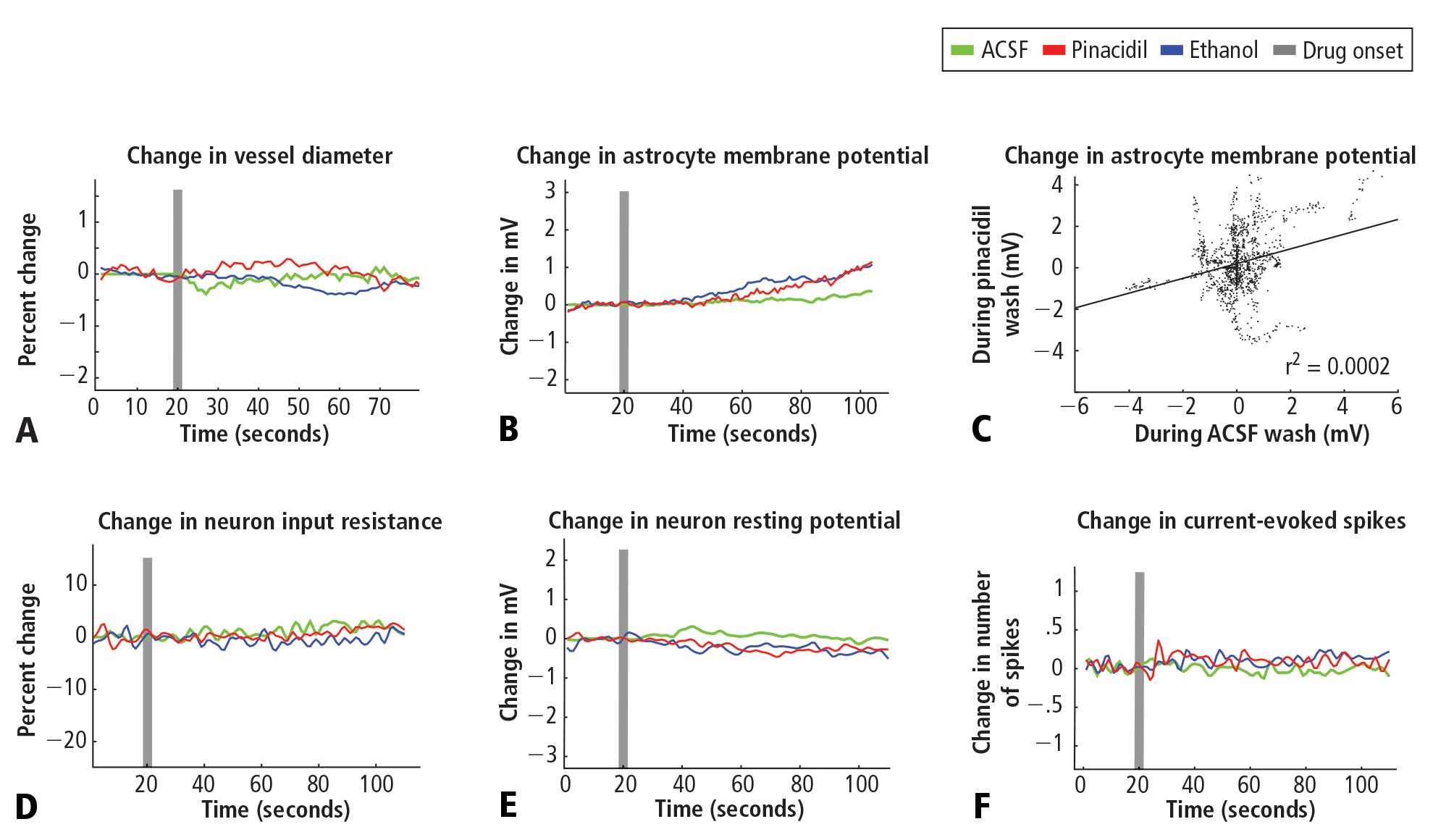
Pinacidil does not impact spiking probability or input resistance in neurons or membrane potential in neurons and astrocytes
In recordings from regular-spiking neurons of pyramidal shape (N = 30), we saw no change in any metric measured. At 50 seconds after application, approximately the time of peak vasodilatory effects in vivo, the resting membrane potential did not change (variation of −0.5 ± 0.9 mV standard deviation; Figure 2E), the spike rate induced by current injection did not change (variation of 0.1 ± 1.1 spikes/stimulation; Figure 2F), and input resistance did not change (variation of −0.7 ± 6.5 MΩ; Figure 2D). Similarly, in a limited subset of recordings from fast-spiking interneurons (N = 3), we did not observe any impact of pinacidil application. All significance tests were paired t tests (P > .10).
Astrocytes (N = 35) also showed no significant effects of pinacidil application, demonstrating only a slow depolarization during application of ethanol (1.3 ± 1.7 mV at 50 seconds after drug application) and pinacidil with ethanol (0.5 ± 2.1 mV) (Figure 2B). When we plotted the change in membrane potential at 10-second intervals since drug application (0 to 80 seconds post-drug), we found no trends in astrocytic response to either ACSF or pinacidil (Figure 2C).
During recordings at double our typical application dose (800 μM), we observed 2 pyramidal cells (out of 8) that showed depolarization (peak of 10 to 15 mV) and a loss of spike initiation capability. Following washout, these cells recovered membrane potential but spiking responses to current injection remained impaired. We did not evaluate the impact of this dose on vascular tone or rhythmic vasomotion.
In contrast to the absence of a detectable impact of pinacidil, we found that the membrane potentials of neurons and astrocytes were sensitive to flow rate. Decreasing flow rate caused a consistent depolarization of up to approximately 10 mV that showed an immediate onset, reaching a new baseline within 2 to 5 seconds; increasing flow rate had the opposite effect. In preliminary experiments, we observed two astrocytes that depolarized on switching to the pinacidil solution. These two recordings were obtained prior to placement of an inline pressure meter in the flow pathway that allowed us to monitor and exclude trials that showed flow changes. In the 35 subsequent recordings that did not have flow changes, we never observed a detectable impact of either ethanol or pinacidil on astroctyes or neurons. We also noted that neurons and astrocytes were more likely to die and/or to lose recording quality during a cycle of ethanol or pinacidil presentation, as opposed to ACSF presentation.